| Home | Conferences | Articles | Guide | Search | |
|
|
Photosensitization of Red Blood Cell Hemolysis: A Brief Review Leonard I. Grossweiner Wenske Laser Center Ravenswood Hospital Medical Center Chicago, IL 60640 USA e-mail: lgflash@aol.com alternate e-mail ligross@bio-laser.org FAX: 773-878-2649
Submitted: Accepted: 16 Sep 199 About the author: Leonard I. Grossweiner is a professor emeritus in physics at the Illinois Institute of Technology and the research director of the Wenske Laser Center at Ravenswood Hospital Medical Center.He is the vice-president of the Midwest Bio-Laser Institute (http://www.bio-laser.org). Professor Grossweiner's research emphasizes medical applications of photosensitization including photodynamic therapy.
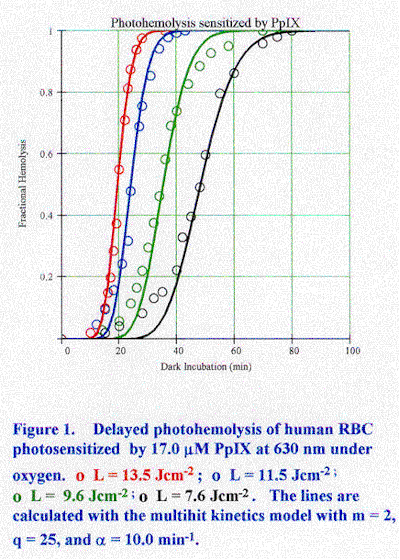
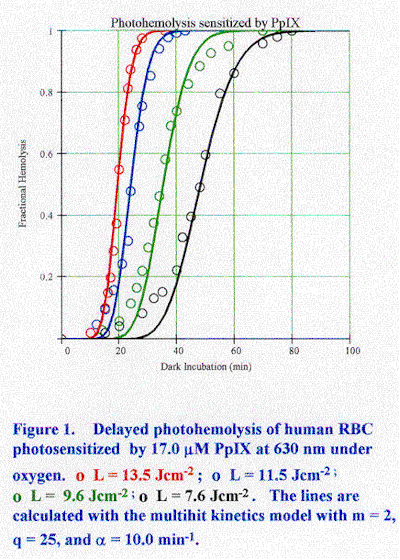
Abstract Photosensitized photohemolysis of red blood cells in vitro has been investigated for many dyes, pigments, and drugs for more than fifty years. Considerable evidence supports the "colloid-osmotic" model which postulates that photochemical damage to the cell membrane leads to cation release and cell swelling followed by hemolysis during dark incubation. The involvement of singlet molecular oxygen has been shown for halogenated fluoresceins, porphyrins, and phthalocyanine photosensitizers. The relative importance of membrane protein and lipid targets has not been resolved. Two recent kinetics models are reviewed which are consistent with the key experimental observations. 1. Introduction It has been known for a long time that exposure of grazing animals to sunlight after they ingest certain plants induces inflammation, ulceration, infection, and in severe cases, convulsions and death. This disease is hypericism [Giese, 1971]. In some regions of Arabia white horses were not used and even unpigmented patches on colored horses were darkened with henna or tobacco juice. In the Tarentine fields of Italy only black sheep were grown because the white ones grew ill after exposure to sunlight. The connection between plants of the genus Hypericum and the sunlight sensitivity of range animals was made in the 19th century, if not earlier. The active photosensitizer was identified with the anthraquinoid pigment hypericin (1,3,6,8,13-hexahydroxy-10,11,dimethyl-phenanthrol [1,10,9,8-opqra] perylene-7,14-dione). Hypericin (HY) is a potent in vitro photosensitizer of red blood cells (RBC). Incubation of an RBC suspension with HY and exposure to light induces membrane lysis and release of hemoglobin to the external medium. This phenomenon is photohemolysis. The early study of Blum [1941] showed that photosensitization of hemolysis by HY requires oxygen. This reaction is a classical type of "photodynamic action" - a harmful biological effect driven by light which requires the presence of a photosensitizer and oxygen. Many chemicals, dyes, and drugs photosensitize in vitro hemolysis. Some of these agents also photosensitize human and animal skin to sunlight. This correlation has motivated the use of photohemolysis as a test for potential skin photosensitizers. The detailed action mechanism of photohemolysis has not been adequately explained notwithstanding more than fifty years of investigations. RBC are highly reproducible model systems for the study of cell membrane damage. There has been considerable recent interest in the heightened sensitivity of cells to oxidant damage in hereditary disorders, such as sickle cell disease and other hemoglobinopathies. Cumulative oxidative damage generates immunologically recognized alterations in the cell surface that may promote binding of autologous antibodies and clearance of senescent cells from the circulation. Photohemolysis may be involved in the action mechanism of photodynamic therapy (PDT) in which occulsion of small blood vessels occurs during the early stages of the procedure [Ben-Hur et al., 1990].The "colloid-osmotic" model is generally accepted as the basic explanation for photohemolysis [Davson and Ponder, 1940]. According to this theory, photochemical damage to the RBC membrane initiates cation leakage, followed by swelling of the cells and hemolysis. These responses to light have been observed experimentally. Other effects which may be related to the photohemolysis mechanism include changes in RBC morphology and fragility induced by dark binding of some photosensitizers, photoperoxidation of membrane lipids, photolysis and photocrosslinking of membrane proteins, and photooxidation of encapsulated hemoglobin. The relationship between specific damage modalities in the photochemical stage of the process and hemolysis during dark incubation has not been established. Different primary photochemical reactions are feasible depending on the photosensitizer and reaction conditions. These include the well-known Type 1 (sensitizer-substrate) and Type 2 (sensitizer-oxygen) photosensitization pathways mediated by the sensitizer triplet state and singlet molecular oxygen (1Dg), respectively. Another possible mechanism involves membrane disruption induced by photolysis products. This paper reviews the current status of photohemolysis research. An attempt has been made to focus on the mechanistic issues by citing illustrative results. The author welcomes additional input and comments.2. Photohemolysis measurements Photohemolysis is observed by exposing an RBC suspension to light and then spinning down the intact cells leaving hemoglobin in the supernatant. A convenient assay procedure utilizes light scattering because the transmission of far-red light by an RBC suspension is proportional to the concentration of intact cells. The photohemolysis rate may be quantified by either of two techniques. In continuous photohemolysis (CPH) the fraction of hemolyzed cells is measured during the irradiation. In post-irradiation or delayed photohemolysis (DPH) the cells are exposed to a brief, non-hemolyzing irradiation and hemolysis is measured during incubation in the dark. DPH and CPH curves are sigmoidal frequently with an initial delay period. The time required to achieve 50% hemolysis (t50) is a useful rate parameter. t50 is measured from the start of irradiation for CPH and from the start of dark incubation for DPH. Photohemolysis kinetics are characterized by a non-linear dependence of the rate on the incident fluence and sensititizer concentration. Values of t50 from less than a minute to many days have been observed. Osmotic fragility is an alternative probe of light-activated damage to RBC membranes. Damaged cells undergo hemolysis at higher bath osmolalities than normal cells. DPH measurements can be performed in two ways. In one procedure, t50 is measured at different values of the incident fluence (F) for a constant total sensitizer concentration (Ct). The data for many sensitizers obey the empirical relation: 1/t50 = aF k ……………………………………………………….………….( 1)where a depends on Ct and k is a constant. Alternatively, the sensitizer concentration can be varied at constant incident fluence. This data correlate with: 1/t50 = bCt j ……………………………………………………………………(2) where b is a function of F and j is a constant. Ideally, the a should have the power dependence j and b should have the power dependence k. Only Ct is varied in CPH measurements leading to: 1/t50 = gCt j '…………………………………………………………………. (3)where function g depends on F and j' may not equal j. Some photosensitizers led to k . 2 and j . 2 (Table 1). This result implies that the photohemolysis rate varied with the square of the absorbed fluence. Other photosensitizers led to k . 2 and j . 1.2-1.4. Lower values of j are considered as anomalies. CPH measurements led to j' < 1, typically 0.5-0.7. The relative steepness (S) of a photohemolysis curve is an additional rate parameter [Al-Akhras and Grossweiner, 1996]. S is defined as t70 /t30, where t70 and t30 are the dark incubation time (in DPH) and irradiation time (in CPH) required for 70% lysis and 30% lysis, respectively. S is not the slope of the steep part of a sigmoidal hemolysis curve when there is a delay period prior to the onset of hemolysis. S has been found to be constant for sets of DPH and CPH curves in which F or Ct were varied (Table 1). Figure 1 shows a typical set of DPH curves for human RBC irradiated at 630 nm after incubation in 17 mM protoporphyrin IX (PpIX) after removal of the unbound sensitizer. The lines are calculated with the multihit kinetics model described in Appendix A. The rate parameters for this data set are k = 1.67 and S = 1.24.2.1. Photosensitization of hemolysis by halogenated fluorescein dyes Cook and Blum [1959] investigated DPH kinetics for human RBC photosensitized by rose bengal (tetraiodotetrachlorofluorescein, RB). The data were plotted as the integral of the gaussian error function and the rate was calculated from as the slope of the hemolysis curves when drawn on a probit plot. From this analysis they deduced that the DPH rate for short irradiations is proportional to the approximate square of the incident fluence and . 1.4 power of the dye concentration. The power dependence on fluence approached 2/3 for long irradiations. These results parallel k, j, and j' in Eq. (1) - Eq. (3). In subsequent work with phloxine B (tetrabromotetrachlorofluorescein, PhB) data fitting to Eq. (1) and Eq. (2) led to k . 2 and j . 1.4. [Valenzeno and Pooler, 1982; Valenzeno, 1984]. Measurements of PhB binding to RBC ghosts showed that the Freundlich absorption isotherm is obeyed according to:Cb = KCt x …………………………………………………………………….. (4)where Cb and Ct t are the bound and total sensitizer concentrations, respectively, and x = 0.61. Thus, if Eq. (4) is substituted in Eq. (2) the effective value of j is close to 2 for bound dye. This result implies that free PhB in the external bath does not induce photohemolysis and supports the square dependence of the DPH rate on the absorbed fluence at low Cb. Relative DPH measurements with ten halogenated fluorescein dyes identified the following factors as related to photosensitivity: (1) the distribution of the sensitizer into a low polarity region of the cell membrane, (2) the triplet state quantum yield, (3) the rate of sensitizer photobleaching [Valenzeno and Pooler, 1982]. The least effective dye was uranin (sodium fluorescein) and the most effective was RB, with PhB in-between. Pooler and Girotti [1986] found that eosin Y isothiocyante (EYNCS) is 50-100 times more effective than eosin Y (tetrabromofluorescein, EY). This dye covalently binds the band 3 (anion channel) membrane protein, which was suggested as a key target for 1Dg . Pooler [1986] proposed that dimers of the band 3 protein are the dual targets responsible for k . 2. Using target theory he showed that this result implies that both members of a dimer pair must be hit at least once. However, measurements with EYNCS led to k . 1.4-1.5 instead of 2 as found for eosin. The discrepancy was attributed to more rapid photobleaching of EYNCS.2.2. Photosensitization of hemolysis by porphyrin derivatives Protoporphyrin IX (PpIX) is the most studied photosensitizer of hemolysis. This interest originated in connection with the human genetic disease erythropoietic protoporphyria (EPP) in which excess PpIX accumulates in the circulation. However, EPP patients have no abnormal hemolysis, although their RBC ex vivo undergo rapid photohemolysis in albumin-free suspensions. Brun and Sandberg [1985] proposed that light exposure induces a transfer of PpIX from hemoglobin to the cell membrane where hemolysis is photosensitized. They suggest that dark and light-activated transfer of PpIX from hemoglobin to the cell membrane and then to albumin in the serum protects EPP patients from in vivo photohemolysis. Photobleaching of PpIX and dark interactions of PpIX with RBC membranes which may affect the results were not considered. DPH measurements on human and bovine RBC with monochromatic 630 nm light led to k . 2 at low PpIX concentrations which decreased slowly to k . 1.5 at high concentrations [Grossweiner et al., 1998]. However, j ranged from . 3 at low concentrations to < 2 at high concentrations. Binding measurements for PpIX showed that x in Eq. (4) is 0.94 for human RBC. The ad hoc correction to j for unbound dye cannot be employed for PpIX as was done for PhB because essentially all PpIX was bound to RBC during the measurements. A factor that was not considered in the earlier work involves the effect of dark PpIX binding on RBC properties, which could have a large influence on j because a different range of photosensitizer concentrations was used for each measurement of t50 . The coefficient a Eq. (1) varied with Ct 1.1 and b in Eq. (2) varied with F3.5. These exponents are not the same as j and k, respectively. Accordingly, the results obtained with monochromatic light show that, although the power functions of Eq. (1) and Eq. (2) are applicable for PpIX, the quantitative predictions of these empirical relations are not supported.A Type 2 photosensitization pathway is favored for PpIX by the high yield of 1 . These exponents are not the same as j and k, respectively. Accordingly, the results obtained with monochromatic light show that, although the power functions of Eq. (1) and Eq. (2) are applicable for PpIX, the quantitative predictions of these empirical relations are not supported.A Type 2 photosensitization pathway is favored for PpIX by the high yield of 1 Dg , higher solubility of oxygen in lipid membranes compared to water, and the longer 1Dg lifetime in hydrophobic media. The effects of additives on photohemolysis provide indirect information about the primary photosensitization mechanisms. Inhibition of DPH by azide ion, tryptophan and 1,4-diazabicyclo[2.2.2]-octane (DABCO) are indirect evidence for the involvement of 1Dg [Grossweiner et al., 1998]. The membrane targets involved in photohemolysis have been the subject of much controversy. The key role of the anion channel protein is supported by studies showing that photosensitization by PpIX affected the rate of carrier-mediated transport [Dubbelman et al., 1980; 1981; 1982; Verweij et al., 1981]. Other evidence supports the involvement of lipid peroxidation. Irradiation of RBC under argon after incubation in PpIX totally inhibited photohemolysis but did not inhibit all protein cross-linking [Lamola and Doleiden, 1980]. The absence of osmotic swelling is a simplifying feature of studies with resealed RBC membranes (ghosts). Irradiation of ghosts incubated with PpIX led to lipid peroxidation and membrane rupture. [Girotti et al., 1985; Grossweiner et al., 1998]. The role of lipid peroxidation is supported also by studies showing that the early stages of PpIX-photosensitized hemolysis altered the fluorescence isotropy of lipophilic probes, while probes bound to membrane proteins were unchanged [Tozzi-Ciancarelli et al., 1989]. The involvement of 1Dg is indicated for hematoporphyrin (HP) by protection with azide ion and faster hemolysis in 98.8% D2O [Sorota et al., 1988]. However, the 1Dg quenchers imidazole and tryptophan were only slightly protective and ascorbate was slightly sensitizing. The PDT drug hematoporphyrin derivative (HPD) is a porphyrin mixture. Irradiation of the higher molecular-weight tumor-localizing fraction of HPD (presumably "dihematoporphyrin ether") in ghost membranes led to the 1268 nm emission of 1Dg and production of membrane-bound 3b-hydroxy-5a-cholest-6-ene-hydroperoxide, which is a unique reaction product of cholesterol with 1Dg [Thomas et al., 1987]. Photosensitization of DPH by chlorin e6 (Chl e6) was inhibited by azide and the antioxidant butylated hydroxytoluene (BHT), while superoxide dismutase (SOD), ethanol, and mannitol had no effect [Frolov and Gurinovich, 1992]. Azide and tryptophan were protective for HP, PpIX, and the chlorin PDT agent, benzoporphyrin derivative monoacid ring A (BPD-MA) [Khalili and Grossweiner, 1997]. Overall, the additive experiments support the Type 2 pathway for porphyrins, although some agents such as ascorbate may have complex interactions with the membranes which inhibit their protective action.2.3. Other photosensitizers and related studies Chemical oxidation of RBC affects their dark and light-induced properties. Treatments of RBC with H2O2 induced shape changes from discoid to echinocyte, lipid peroxidation, transfer of inner membrane lipids to the cytosol, increased cation permeability, complexation of hemoglobin to spectrin, oxidation of hemoglobin, and decreased membrane deformability [Williams et al., 1995; Van der Zee et al., 1985; Snyder et al., 1988; Chen et al., 1991; Lagerberg et al., 1996]. Oxidation of hemoglobin in H2O2 treated cells was accompanied by greatly increased adherence of hemoglobin to the membrane [Chen et al., 1991]. Prior PpIX binding protected against lipid peroxidation by H2O2 [Williams et al., 1995]. Furthermore, RBC which had been treated with H2O2, washed, and then incubated with PpIX could not be lysed by light [Grosssweiner et al., 1998]. Phthalocyanine sulfonates are photosensitizers of DPH. The most effective central cation was aluminum followed by zinc and the metal-free derivative; the paramagnetic derivatives with copper, iron, and cobalt were inactive [Sononda et al., 1987]. Mediation by 1Dg was shown for aluminum phthalocyanine tetrasulfonate (AlPcS) by the enhanced DPH rate in 98% deuterated media and protection by tryptophan [Ben-Hur et al., 1994]. The DPH rate for chloroaluminum phthalocyanine sulfonate (AlPcSCl) was inhibited by azide, increased by ascorbate, and unaffected by mannitol and SOD [Rosenthal and Ben-Hur, 1992]. The totality of results for porphyrin derivatives supports the key role of 1Dg in the photohemolysis mechanism, although contributions from other pathways cannot be ruled out. Recent measurements of the 1Dg quantum yield for some of these compounds in pH 7.4 phosphate buffer plus 1% Triton X-100 are: HP, 0.73; PpIX, 0.56); Photofrin7, 0.91; Chl e6, 0.64; BPD-MA, 0.84; AlPcS, 0.38 [Fernandez et al., 1997].Measurements with HY showed that imidazole was only slightly protective while ascorbic acid promoted CPH [Senthil et al., 1994]. However, photosensitization of lipid peroxidation in RBC ghosts membranes by HY was strongly protected by the 1 Dg quenchers azide, imidazole, and DABCO [Senthil et al., 1992]. The furocoumarin derivatives employed for "psoralen-plus-UVA" (PUVA) photochemotherapy of human skin diseases are primarily Type 1 photosensitizers of DNA. However, some psoralens are Type 2 photosensitizers of RBC via the 1Dg pathway including psoralen, 8-methoxypsoralen, 3-carbethoxypsoralen, and angelicin [Dall'Acqua et al., 1987; Potepenko, 1991]. Results for some photosensitizers of hemolysis do not indicate the major involvement of 1Dg . Photosensitizers which are active under oxygenated and anoxic conditions include a-alkylamino-2-arylquinolinemethanol anti-malarial compounds [Epling and Sibley, 1987], the non-steroidal anti-inflammatory drug carprofen [De Guidi et al., 1993], and the antifungal drug griseofulvin [Fujita et al., 1993]. Polytriptyline and chlorpromazine generate membrane-bound photoproducts that promote RBC lysis [Gasparro and Kochever, 1982; Kochever and Hom, 1983].Direct evidence for cation release and cell swelling have been reported in conjunction with photohemolysis studies. Irradiation of human RBC in the presence of PpIX led to temperature-independent K+ leakage which was attributed to photooxidation of amino acid residues in membrane proteins [Dubbleman et al, 1980]. Secondary protein crosslinking was observed in RBC ghosts and in RBC during dark incubation at 37EC after irradiation in the presence of PpIX at 0EC. Measurements with AlPcSCl showed that release of K+ from irradiated human RBC occurred without lipid peroxidation [Deuticke et al., 1989]. The leak permeability was accompanied by the formation of a low number of relatively large pores (0.45-0.75 nm) at < 1 per cell. Direct measurements of cell swelling with AlPcSCl showed that the cell volume increased linearly with the light fluence, after an initial delay, and increased with temperature [Ben-Hur et al., 1990]. The rate of volume increase depended on the cation present in the medium in the order: Rb+ = Cs+ > K+>Na+>Li+. CPH studies with a-terthienyl irradiated with UVA showed that K+ release occurred first, followed by hemolysis and then loss of acetylcholinesterase activity [Wat et al., 1980]. Evidence for a Type 2 pathway in this system is the requirement for oxygen and the formation of membrane-bound 3b-hydroxy-5a-cholest-6-ene-hydroperoxide. The applicability of the colloid-osmotic mechanism for the non-photodynamic agent griseofulvin is indicated by measurements showing that hemolysis is preceded by K+ release [Fujita et al., 1993].3. Photohemolysis kinetics The key feature of photohemolysis is that cell lysis involves light-activated and thermal processes acting in concert. Accordingly, it is difficult to compare the photosensitization rate and energy efficiency for measurements performed under different treatment conditions. A kinetics model for photohemolysis should predict the sigmoidal form of photohemolysis curves and the dependence of the photohemolysis rate on the incident fluence, photosensitizer concentration, and other relevant variables. The model of Pooler postulates the occurrence of specific processes within the general framework of the colloid-osmotic model [Pooler, 1986a; 1986b]. The key assumption is that the light-activated events affect only the cation permeability of the RBC membranes, while the osmotic and lytic behavior of the cells are identical to those of normal cells. It is assumed that the cation permeability follows similar power functions of the fluence and sensitizer concentration as 1/t50 in Eq. (1) and Eq. (2). The relationship between permeability and cell volume is calculated for normal RBC from the model of Freedman and Hoffman [1979] in which the osmotic coefficient of hemoglobin has a non-linear dependence on the interior hemoglobin concentration. Fractional lysis (fL) is then related to the relative cell volume (v) by an empirical expression: fL(v) = 1/[1 + exp[K(v - v50)]], where K is a steepness constant and v50 is the relative cell volume at 50% lysis. Model calculations predict the time-dependence of K+ loss, cell swelling, and hemolysis. Numerical calculations for PhB assuming k = 2 and j = 1.33 are in good agreement with experimental DPH curves. The kinetics model of Grossweiner [Al-Akhras and Grossweiner, 1996] extends the early target theory model of Pooler[1986] by expressing the light-activated and the thermal incubation stages in terms of multihit target theory (Appendix A). The key assumptions are: (1) efficient photohemolysis takes place by thermal expression of pre-lytic photochemical damage; (2) damage thresholds exist for the photochemical and thermal stages of the overall process. A photochemical hit number (m) is defined corresponding to the kinetic order of the light-activated process, i.e., a one-hit process (m = 1) depends linearly on F, a two-hit process (m = 2) depends on L2, etc. A thermal hit number (m) is which is related to the extent of cooperativity in the thermal stage. Typical values of q from model fitting to data are 15-30 [Al-Akhras and Grossweiner, 1996; Khalili and Grossweiner, 1997]. This model predicts that DPH curves obey the functional form of Eq. (1) and Eq. (2) at low fluence with k = j = m and that similar power functions are approximately valid at higher fluence with lower values of the exponents. The experimental results in Table 1 indicate that m = 2 is applicable for HY-Na, BPD-MA, and HP. Possible explanations for the anomalous values of j with the halogenated fluorescein, Chl e6 , and PpIX are discussed above. The multihit analysis for CPH leads to j' =1/2 for m = 1, j' = 2/3 for m = 2, j' = 3/4 for m = 3, etc The prediction j' = 2/3 for m = 2 is consistent with experimental CPH results (Table 1) as well as the dependence of photohemolysis rate on fluence reported by . This model predicts that DPH curves obey the functional form of Eq. (1) and Eq. (2) at low fluence with k = j = m and that similar power functions are approximately valid at higher fluence with lower values of the exponents. The experimental results in Table 1 indicate that m = 2 is applicable for HY-Na, BPD-MA, and HP. Possible explanations for the anomalous values of j with the halogenated fluorescein, Chl e6 , and PpIX are discussed above. The multihit analysis for CPH leads to j' =1/2 for m = 1, j' = 2/3 for m = 2, j' = 3/4 for m = 3, etc The prediction j' = 2/3 for m = 2 is consistent with experimental CPH results (Table 1) as well as the dependence of photohemolysis rate on fluence reported by the halogenated fluorescein, Chl e6 , and PpIX are discussed above. The multihit analysis for CPH leads to j' =1/2 for m = 1, j' = 2/3 for m = 2, j' = 3/4 for m = 3, etc The prediction j' = 2/3 for m = 2 is consistent with experimental CPH results (Table 1) as well as the dependence of photohemolysis rate on fluence reported by Cook and Blum [1959]. Another prediction is that the relative steepness of DPH curves depends only on q. In general, the multihit model predicts sets of DPH and CPH curves for a given photosensitizer in terms of measurable quantities and the same arbitrary parameter which is the coupling constant a. The lines in Fig. 1 for DPH photosensitized by 20 mM PpIX (Cb = 17 mM) were calculated with m = 2 and q = 25, as deduced from the data for k . 2 and S = 1.24, and a = 10.0 min-1 which gave the best overall fit to the data.3. Energetics of Photohemolysis Energy efficiency is a variable quantity for photohemolysis owing to the nonlinear dependence of the rate on the absorbed fluence. However, an apparent energy efficiency can be calculated for measurements made at the same t50 Energy efficiency is a variable quantity for photohemolysis owing to the nonlinear dependence of the rate on the absorbed fluence. However, an apparent energy efficiency can be calculated for measurements made at the same t50. The inverse quantum yield of a photochemical process is the number of photons absorbed for each product molecule generated. An equivalent parameter for photohemolysis is the number of photons absorbed by photosensitizer molecules leading to a givent50 .This quantity can be calculated on a "per sensitizer molecule" basis ( hss) or a "per cell" basis (hcc). hs s parallels the conventional quantum yield for homogeneous systems in which all photosensitizer molecules are assumed to be equivalent. hc is more relevant for those systems in which the unbound sensitizer is inactive. Assuming that all bound sensitizer molecules are equivalent leads to:1/ hc c = 1.928 x 10-5 F0F0lNbea /Nrbc ………………………………………………(5)where F0 (Jcm-2) is the incident fluence, l(nm) is the irradiation wavelength, Nb is the mean number of sensitizer molecules per cell, ea a (M-1cm-1) is the molar absorbance of the bound sensitizer, and Nrbc (cm-3) is the RBC concentration. Values of F0 in Eq. (5) may be calculated from Eq. (1) at the given t50 . Table 2 summarizes the calculations for DPH of bovine and human RBC photosensitized by PpIX at 630 nm [Grossweiner et al., 1998]. The numerical values include additional corrections for saturation of light absorption at high PpIX concentrations and inner filtering of incident light by hemoglobin. The energy efficiency is about ten-fold higher for t50 = 24 h compared to 0.5 h. The same data for human RBC led to hs s . 250 at t50 = 0.5h and hs . 30 at t50 = 24h. Thus, photosensitized hemolysis is a relatively efficient process on a "per sensitizer molecule" basis.4. Discussion and Conclusions The available facts are consistent with the basic tenants of the colloid-osmotic model. However, many details remain unresolved. The weight of evidence indicates that photodynamic agents act by the attack of 1 Dg on membrane transport proteins, leading to K+ release followed by cell swelling. It is not known whether this process alone is responsible for efficient hemolysis during the dark incubation stage. The square dependence of the DPH rate on fluence indicates that a second cellular target must bedamaged, possibly structural membrane proteins or the lipid part of the membrane. There is evidence for these reactions with PpIX. The dark effects of sensitizer binding on membrane properties were not given adequate consideration in earlier work. The sensitizer-RBC complex is a unique system with different photophysical and photochemical properties than native RBC. A valid microscopic mechanism must be consistent with the rate kinetics. Considering the variabilities in the photohemolysis process, the applicability of Eqs. (1)-(3) for so many different sensitizers is quite surprising. However, quantitative measurements with PpIX indicate that these power relations are only semi-quantitative. The same key assumption is made in the kinetics models of Pooler and that of Grossweiner: Photohemolysis takes place by dark expression of photochemical damage. The kinetics model of Grossweiner postulates that the sigmoidal curves result from photochemical and thermal damage thresholds. The superlinear dependence of 1/t50 on fluence clearly implies the existence of a photochemical damage threshold below which hemolysis is absent or highly inefficient. The concept of a thermal damage threshold is more speculative. Sigmoidal photohemolysis curves may result from other factors. One possibility is that RBC in a given population have a distribution of fragilities according to their physiological ages. The multihit model correctly predicts the power-function relations of Eqs. (1)-(3) in the limit of low fluence and the constant relative steepness of sets of DPH and CPH curves without any other assumptions. The model of Pooler emphasizes the effects of membrane alterations on the RBC osmotic properties. This approach permits detailed modeling of K+ release and cell swelling which are consistent with experiments. However, an arbitrary sigmoidal function is assumed as well as the functional forms of Eq. (1) and Eq. (2). In summary, the Pooler model describes the dark incubation stage of the process in detail while employing an empirical expression for the light-activated stage. The multihit model postulates a specific kinetics mechanism for the photochemical stage and arbitrarily extends an equivalent formalism to the dark stage. Taken together the two approaches provide a useful quantitative description of photohemolysis rate kinetics, although arbitrary assumptions leading to the numerical results are employed in both models.Appendix A. Multihit model of photohemolysis rate kinetics The key assumptions of the multihit kinetics model are: (1) Photohemolysis takes place by thermal expression of pre-lytic photochemical damage. (2) Damage thresholds exist for the photochemical and thermal stages of the overall process which can be described in terms of "hits" on a target. The photochemical hit number (m) specifies the kinetics order of the photochemical stage. Thermal "hits" are a hypothetical concept related to the cooperativity of the thermal process. The mathematical implementation of the model is based on multihit target theory. The mean number of photochemical hits on a "per cell" basis (n) after an incident fluence (It') is expressed as: n = It' N bsag It' Nbsag ………………………………………………………………..(A1)where I is the incident photon fluence rate, t' is the DPH irradiation time, N bNb is the average number of sensitizer molecules bound to a red cell, sa a is the photon absorption cross section of bound sensitizer molecules, and g is a number which is proportional to the probability that an absorbed photon leads to a photochemical hit. According to target theory the probability ppm(n) of accumulating at least m hits after an effective light dose n is given by:p m(n) = 1 - exp(-n)S(n)v/v! …………………………………………………(A2)where the summation is from v = 0 to v = m-1. Numerical calculations based on Eq. (A2) show that p pm = 2 at n . m at high m. The key assumption of the thermal stage is that the mean number of thermal "hits" after dark incubation time t depends linearly on ppm. The fractional lysis after dark incubation time t is given by: fq(t) = 1 - exp(- apmtt)S (apmt)u/u! …………………………………….…..(A3)where the summation is from u = 0 to u = q-1 and a a is a constant that couples the photochemical and thermal stages. Equation (A3) indicates that t t50 is fully determined by ppm and a according to:t 50 = DDq/ppma ………………………………………………………………..(A4)where D q Dq is a number close to q. Numerical calculations show that DDq = 4.67 for q = 5, 9.67 for q = 10, 19.67 for q = 20, 24.67 for q = 25, and 29.67 for q = 30, etc . Equation (A3) shows that S depends only on the upper limit q. An important conclusion derived from this model is that DPH curves obey the exponential form of Eq. (1) and Eq. (2) at low effective light dose and k = j = m in this limit. This result is easily shown in the limit of low effective light dose, in which case ppm . nnm/m! and ffq(t) . (a pm tt)q/q! . {In this limit t50 is replaced by a shorter time t0 at which ffq (t) = f0 }.The same approximation shows that the relative steepness S depends only on q. Numerical calculations show that the power dependence of Eq. (1) remains approximately valid at higher effective light dose but with lower values of the exponent k. The modeling results for CPH are obtained by setting the irradiation time t in Eq. (A3) equal to the incubation time t'. Numerical calculations show that j' =1/2 for m = 1, j' = 2/3 for m = 2, j' = 3/4 for m = 3, etc. This result is derived explicitly in the limit of low effective light dose leading to j' = m/(m + 1). References Al-Akhras, M. A. and L. I. Grossweiner (1996) , Sensitization of photohemolysis by hypericin and Photofrin R. J. Photochem. Photobiol. B Biol., 34: 169-175.Beaton, S., (1995), R. A. McPherson, and L. Tilley, Alterations in erythrocyte organization induced by the photosensitizer hematoporphyrin derivative. Photochem. Photobiol., 62: 353-355.Ben-Hur, E. (1990), A. Orenstein, A. Livne, and I. Rosenthal , Photosensitized oxidation of human red blood cells: Cation effects on volume changes and relevance to blood vessel occlusion. Lasers Life Scis., 3: 245-253.H. F. Blum, H. F. (1941) , Photodynamic Action and Diseases Caused by Light, Hafner, New York,Brun, A. and S. Sandberg (1985) , Photodynamic release of protoporphyrin from intact erythrocytes in erythropoietic protoporphyria: The effects of small repertitive light doses. Photochem. Photobiol., 41: 535-541.Chen, M. (1991), M. P. Sorette, DT.-Y. Chiu, and M. R. Clark , Prehemolytic effects of hydrogen peroxide and t-butylhydroperoxide on selected red cell properties. Biochim. Biophys. Acta , 1066: 193-200.Cook, J. S. and H. F. Blum (1959) , Dose relationships and oxygen dependence in ultraviolet and photodynamic hemolysis. J. Cell. Comp. Physiol., 53, 41-60.D'all Acqua, F. (1987), G. Tamborrino, D. Vedaldi, and P. Arslan , Photohemolysis of erythrocytes by furocoumarins. Med. Biol. Environ.,15, 43-46.Davson, H. and E. Ponder (1940) , Photodynamically induced cation permeability and its relation to hemolysis. J. Cell. Comp. Physiol., 15, 67-74.De Guidi , G. (1993), R. Chillemi, S. Giuffrida, and G. Condorelli, Molecular mechanisms of drug photosensitization 4. Photohemolysis sensitized by carprofen. J. Photochem. Photobiol. B Biol., 17, 239-246.Deuticke, B. (1989), U. Henseleit, C. M. Haest, K. B. Hell, and T. M. A. R. Dubbelman , Enhancement of translayer mobility of a membrane liquid probe accompanies formation of membrane leaks during photodynamic treatment of erythrocytes. Biochim. Biophys. Acta, 982, 53-61.Dubbelman, T. M. (1980) A. F. De Goeij, and J. Van Steveninck , Protoporphyrin-induced photodynamic effects on transport processes across the membrane of human erythrocytes. Biochim Biophys Acta, 595, 133-139.Dubbelman, T.M.A.R. (1982), C. Haasnoot, and J.Van Steveninck . Temperature dependence of photodynamic red cell membrane damage. Biochim. Biophys. Acta, 601, 220-227.Verweij, H. (1981), T.M.A.R.Dubbelman, and J. Van Steveninck , J. Photodynamic protein cross-linking. Biochim. Biophys. Acta, 647, 87-94.Dubbelman, T.M. (1981), A. F. De Goeij, K. Christianse, and J. Van Steveninck , Protoporphyrin-induced photodynamic effects on band 3 protein of human erythrocyte membranes. Biochim. Biophys. Acta, 649, 310-316.Emiliani, C. and M. Delmelle (1981) , The lipid solubility of porphyrins modulates their phototoxicity in membrane models. Photochem. Photobiol., 37, 487-490.Epling, G. A. and M. T. Sibley (1987) , Photosensitized lysis of red blood cells by phototoxic antimalarial compounds. Photochem. Photobiol., 46, 39-43.Fernandez, J. M. (1997), M. D. Bilgin, and L. I. Grossweiner , Singlet oxygen generation by photodynamic agents, J. Photochem. Photobiol. B: Biol,. 37, 131-140.Freedman, J. C. (1979) and J. F. Hoffman , Ionic and osmotic equilibria of human red blood cells treated with nystatin. J. Gen. Physiol., 74, 157-185.Frolov, A. A. (1992) and G. P. Gurinovich , The laws of delayed photohaemolysis sensitized by chlorin e6. J. Photochem. Photobiol. B: Biol., 13 , 39-50.Fujita, H. (1993), N. Inukai, and I. Matsuo, In vitro photosensitized lysis of red blood cells by an antifungal drug griseofulvin. J. Photochem. Photobiol. B: Biol., 17 , 77-80.Gasparro, F. P and I. E. Kochever (1982) , Investigation of protriptyline photoproducts which cause cell membrane disruption. Photochem. Photobiol., 35, 351-358.Giese, A.C. (1971) , Photosensitization by natural pigments. In A. C. Giese (ed) Photophysiology, Vol 6, New York, Academic Press, pp. 77-129.Girotti, A.W. (1985), J. P. Thomas and J. E. Jordan , Lipid peroxidation in erythrocyte ghosts: sensitization of the membranes toward ascorbate and superoxide-induced peroxidation and lysis. Arch. Biochem. Biophys., 236, 238-251.Grossweiner, L. I. (1998), J. M. Fernandez, and M. D. Bilgin , Photosensitisation of red blood cell haemolysis by photodynamic agents. Lasers Med. Sci., 13, 42-54,Khalili, M. and L. I. Grossweiner (1997) , Sensitization of photohemolysis by benzoporphyrin derivative monoacid ring A and porphyrins. J. Photochem. Photobiol. B: Biol., 37, 236-244.Kochevar, I. E and J. Hom. (1983) , Photoproducts of chlorpromazine which cause red blood cell lysis. Photochem. Photobiol., 37, 163-168.Lagerberg, J.W.M.(1996), M. Williams, A.C.E. Moor, et al. , The influence of merocyanine 540 and protoporphyrin on physicochemical properties of the erythrocyte membrane. Bichem. Biophys. Acta 1728, 247-253.Lamola, A. A. and F.H. Doleiden (1980) , Cross-linking of membrane proteins and protoporphyrin-sensitized photohemolysis. Photochem. Photobiol., 31, 597-601.Pooler, J. P. (1985) , A new hypthesis for the target in photohemolysis: Dimers of the band 3 protein. Photochem. Photobiol., 43, 263-266.Pooler, J. P. (1985a) , The kinetics of colloid osmotic hemolysis. II. Photohemolysis, Biochim. Biophys. Acta., 812, 199-205.Pooler, J. P. (1985b) , The kinetics of colloid osmotic hemolysis. I. Nystatin-induced lysis, Biochim. Biophys. Acta, 812, 193-198.Pooler, J. P. and A. W. Girotti (1986) , Photohemolysis of human erythrocytes labeled in band 3 with eosin-isothiocyanate, Photochem. Photobiol., 44 , 495-499Potapenko, A. Ya (1991) , Mechanisms of photodynamic effects of furocoumarins, J. Photochem. Photobiol. B: Biol., 9, 1-32.Potapenko, A. Ya (1986), S. Wunderlich, F. Pliquett, et al. , Photosensitized modification of erythrocyte membranes by furocoumarins. Photobiochem. Photobiophys., 10, 175-180.Rosenthal, I. and E. Ben-Hur (1992) , Ascorbate-assisted phthalocyanine-sensitized photohaemolysis of human erythrocytes. Int. J. Radiat. Biol., 62, 481-486.Senthil, V. (1994), L. R. Jones, K. Senthil, and L. I. Grossweiner , Hypericin photosensitization in aqueous model systems, Photochem. Photobiol., 59, 40-47.Senthil, V. (1992), J. W. Longworth, C.A. Ghiron, and L. I. Grossweiner, Photosensitization of aqueous model systems with hypericin. Biochim. Biophys. Acta, 1115, 192-200.Snyder, L.M. (1988), N. L. Fortier, L. Leb, et al. The role of membrane protein sulfhydryl groups in hydrogen-peroxide-mediated membrane damage in human erythrocytes. Biochim. Biophys. Acta, 937, 229-240.Sonada, M. (1987), C. Murali Krishna, and P. Riesz , The role of singlet oxygen in the photohemolysis of red blood cells sensitized by phthalocyanine sulfonates, Photochem. Photobiol., 46, 625-631.Tozzi-Ciancarelli, M.G.(1989), A. Di Giulio, E. Troiani, et al ., Human erythrocyte damage at the initial stages of oxidative stress. Cell Biophys., 3, 225-234.Valenzeno, D. P. and J. P. Pooler (1982) , The concentration and fluence dependence of delayed photohemolysis. Photochem. Photobiol., 35, 427-429.Valenzeno, D.P. (1984) , Photohemolytic lesions: Stoichiometry of creation by phloxine B. Photochem. Photobiol., 40, 681-688.Van der Zee, J. (1985), T. M. A .R. Dubbelman, and J. Van Steveninck , Peroxide-induced membrane damage in human erythrocytes. Biochim. Biophys. Acta, 818, 38-44.Wat, C.K. (1980), W. D. MacRae, E. Yamamoto, et al. , Phototoxic effects of naturally occurring polyacetylenes and a-terthienyl on human erythrocytes. Photohem. Photobiol., 32, 167-182.Williams, M. (1995), J. W. M. Lagerberg, J. Van Steveninck, and J. Van der Zee, The effect of protoporphyrin on the susceptibility of human erythrocytes to oxidative stress: exposure to hydrogen peroxide. Biochim. Biophys. Acta, 1236, 81-88.Table 1 |
{kind=link}
{kind=link}
{kind=link}