| Site home page | Conference home page | Discussion |
SOME GENERAL REGULARITIES OF IONIZING AND
633 nm LASER RADIATION ACTION ON BACTERIA
K. SH. VOSKANYAN
Laboratory of Nuclear Problems,
141980 Dubna, Russia
It is well known, that 633 nm laser radiation has the wide spread use in biology and medicine (1,2). Therefore the elucidation of mechanisms and regularities of these radiations action on cells is of a big interest. It seems, that the search of general regularities of laser and ionizing radiations action on cells is one of the possible ways for the clarification of laser radiation influence on cells.
With this aim, the action of the 633 nm laser and ionizing (x-rays, g-rays and a-particles) radiations on E.coli bacteria cells of different genotypes has been investigated.
MATERIAL AND METHODS
Isogenic strains of E.coli K-12 were used: wide type - AB 1157, sensitive mutants AB 2463 (rec A13-), P 3478 (pol Al-), super resistant mutant BL 1114 (Gamr 444) and strain Hfr H. CW helium-neon lasers LG-52-3 and LG-75, l= 633 nm (radiation power 2 mW and 4,8 mW accordingly) has been exploited. Irradiation by a-particles was performed on flat source 239Pu, with the dose power 21 Gy/min, average value of LET 100 keV/mm. We also used 137Cs ( g- rays) with dose rate 35 Gy/min and unfiltered X-rays (apparatus RUP-200-20-5, voltage 200 kV) with the same dose rate.
Before irradiation cells were grown on solid complete nutrient medium PPA (plan-peptone agar from Gamaleya Institute of Microbiology, Moscow, Russia) for 24 hours at 370C. Irradiation of cells was performed on solid agar (4% agar) without nutrients, in a monolayer, at room temperature. Under successive cell irradiation by ionizing and laser radiations, the time interval between two kinds of irradiations never exceeded 120 sec.
Cells survival was defined by macrocolony calculation that grows after 48-hours at 370C.
To express the mutated cells we applied the "sandwich seeded" method (3): irradiated samples were washed from the pieces of solid agar with physiological solution and were seeded in Petri dishes with 12 ml PPA (in which we added 50 ml threpheniltetrazoliumchloride before autoclaving, and 50 ml of 20% lactose, one of the selective medium after it) and then we also filled in 12 ml of the same medium. Mutated colonies were red. The counting of the mutant colonies was carried out with the help of a MBC-9 microscope.
The mutation frequency was measured as a ratio of mutant clone (Nm) to the number of surviving cells (N). The number of spontaneous mutations determined by this technique in control (nonirradiated) culture was equal to 10-7 cells in the average.
5-7 independent experiments were performed and the standard errors for each experimental point were less than 5%.
RESULTS AND DISCUSSION
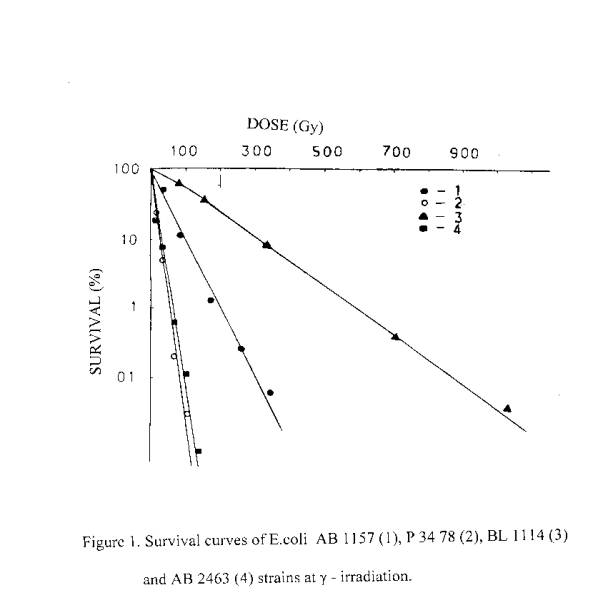
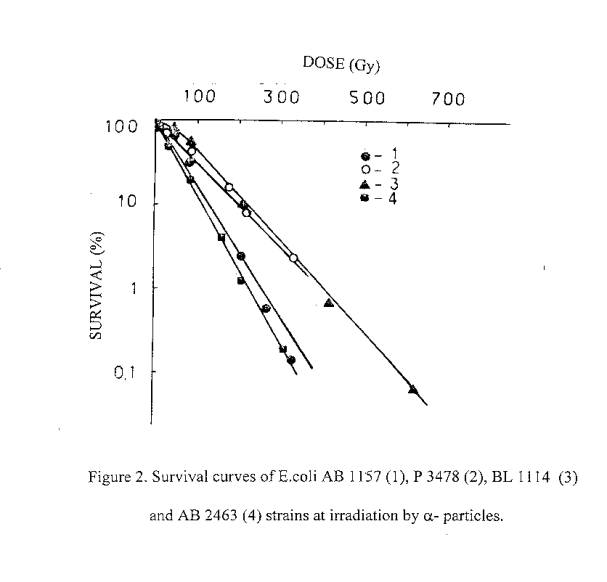
It is well known, that a lethal action of the ionizing radiation in dependence on dose is a reality for all organisms. Therefore, it was a subject of interest to investigate the possible lethal action of laser radiation. We have performed a serial of experiments to investigate the lethal action of the ionizing and laser radiations on bacteria. It was found out, that a lethal action of laser radiation similar to ionizing, takes place in dependence on dose. Obtained results are presented in figures 1-3. As can be seen, the sensitivity of E.coli strains to the laser and rarely ionizing radiations is described by similar in directions rows. Gamr mutant cells are most radioresistant, poIA- and recA- mutants - least radioresistant, and <<wide>> type cells are intermediate. Under the exposure to a-particles, the difference between the sensitivity of various genotypes is decreases due to the high LET of a - particles.
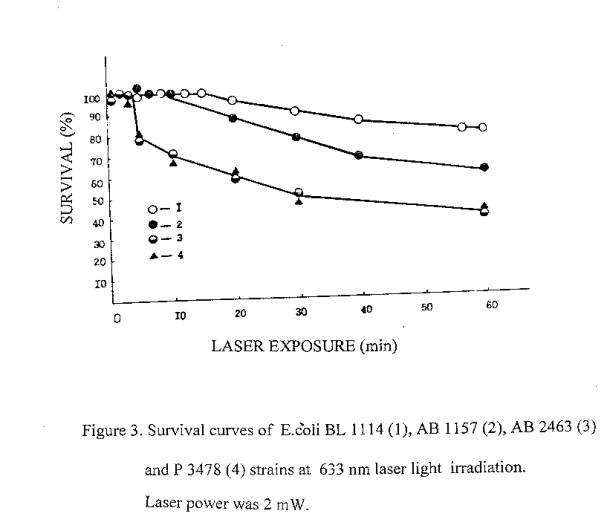
The dose dependence on biological effects is generally used for quantitative evaluations of radiations biological action. However, in some cases the effect of exposure depends also on the dose power (4). This dependence is characterized in biology by the conception “time factor” (5). Observed decreasing of irradiation effectiveness at low dose powers usually considered as a result of cells recovery systems activity (6). Decreasing of the irradiation effectiveness for low radiation powers commonly registrated in the cases, when the quantity of investigated powers are vary not less than in one order, at the conditions, which are not hinder cells recovery.
For the elucidation of the role of "time factor" in biological efficiency of laser radiation we investigated the dependence of E.coli bacteria cells survival on exposure time of various laser radiation power. The results presented in figure 4 testify that the effect initiated by given dose is defined only by its quantity: the intensity of the radiation is essentially immaterial. As an example, one can see that 40 min laser exposure at 1,9 mW laser power and 16 min laser exposure at 4,8 mW laser power caused to similar effect - cells survival decreased to 30%. The density of energy in both cases is equal to 6,49 105 J/m2. Similar picture has been observed under all other values of cells survival.
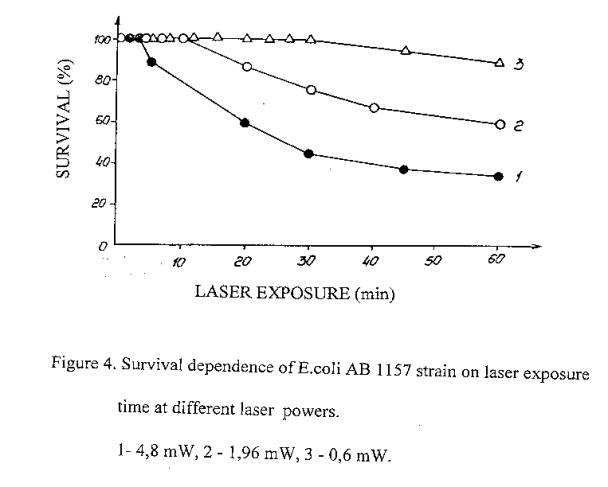
Hence, over the examined laser radiation powers interval, which are varied less than in one order, irradiation efficiency is independent on dose power, as in the case of ionizing radiation.
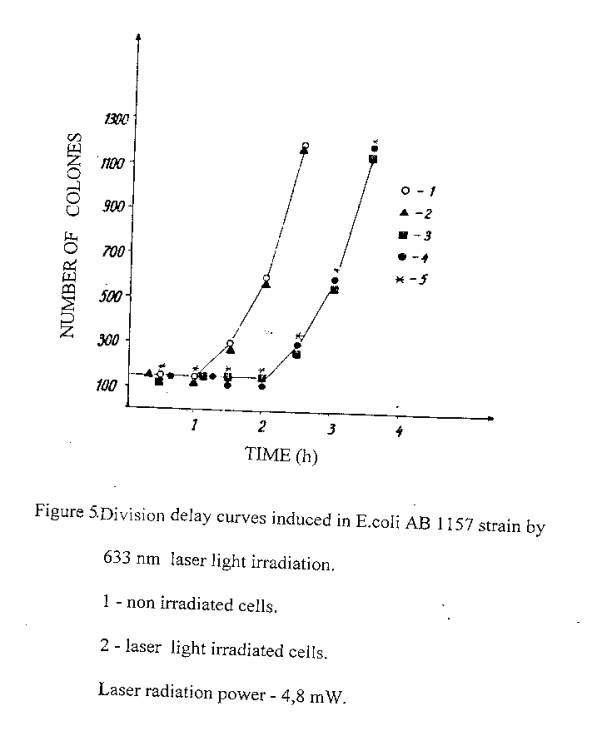
It has been known another general effect of ionizing irradiation - temporal suppress of cell division (7). Our experiments were also directed to clarify this effect in the case of the laser radiation action (5). To registrate the cells division time, the control and irradiated cells were seeded on the nuclear filters (holes diameter - 0,53 mm), which were coated on a solid complete nutrient medium and located in thermostat at 370C. Keeping them for fixed time in such conditions, cells were washed from filters and then seeded on solid agar in Petri dishes. Data on division was carried out by increases number of colony - forming cells. It is seen from figure 5, that the control and 5 sec laser exposed cells were divided after 1,5 hours, while prolonged laser exposed cells (12 sec, I and 2 min) were divided in 2,5 hours.
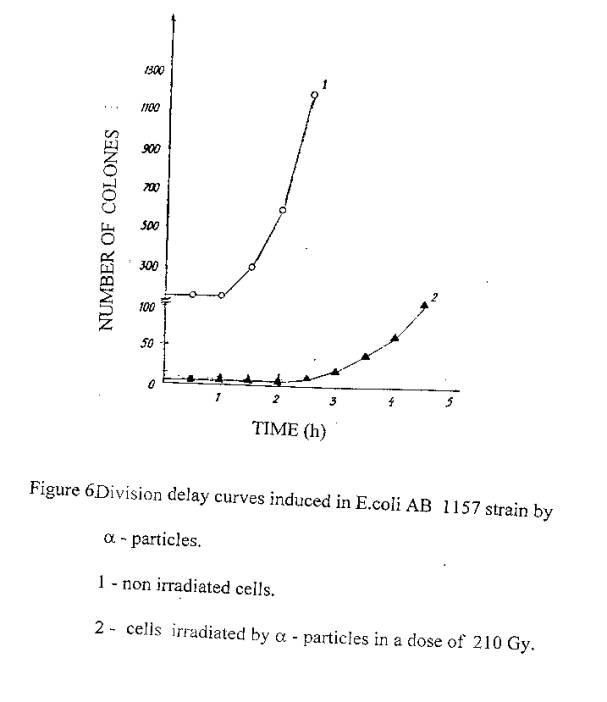
In figure 6 are presented data on similar experiments with cells irradiated in a dose of 210 Gy by a- particles. The division time of this cells was also 2,5 hours.
Thus, cells exposure to laser radiation at 633 nm, similar to ionizing irradiation, causes the suppress of cells division.
On the basis of our previous findings on the lethal effects of 633 nm laser irradiation on E.coli it is expected that laser induced cell inactivation is due to the induction of DNA damages (similar to ionizing radiation). Since ionizing radiation gives rise to lethal effects accompanied by mutations due to the induction of DNA damage, we also tested the induction of mutations by red light in order to show the possible induction of damage to DNA. With this aim experiments were carried out to find out the mutagenic action of 633 nm laser radiation on bacteria cells. The induction of Lac- mutations in E.coli Hfr H has been investigated. Experimental values of mutagenic and lethal effectiveness of 633 nm laser radiation and ionizing radiation are shown in figures 7 and 8. It is obvious from the figures that in both cases irradiation had mutagenic effects on bacteria.
On the basis of our previous findings on the lethal effects of 633 nm laser irradiation on E.coli it is expected that laser induced cell inactivation is due to the induction of DNA damages (similar to ionizing radiation). Since ionizing radiation gives rise to lethal effects accompanied by mutations due to the induction of DNA damage, we also tested the induction of mutations by red light in order to show the possible induction of damage to DNA. With this aim experiments were carried out to find out the mutagenic action of 633 nm laser radiation on bacteria cells. The induction of Lac- mutations in E.coli Hfr H has been investigated. Experimental values of mutagenic and lethal effectiveness of 633 nm laser radiation and ionizing radiation are shown in figures 7 and 8. It is obvious from the figures that in both cases irradiation had mutagenic effects on bacteria.
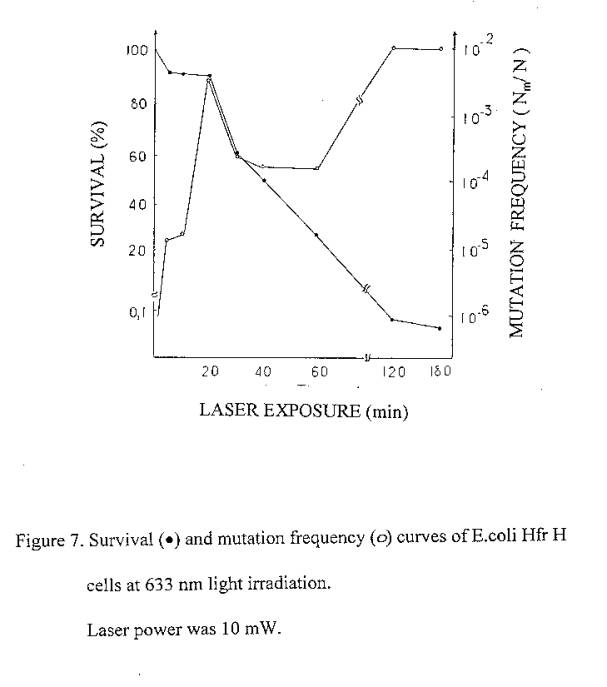
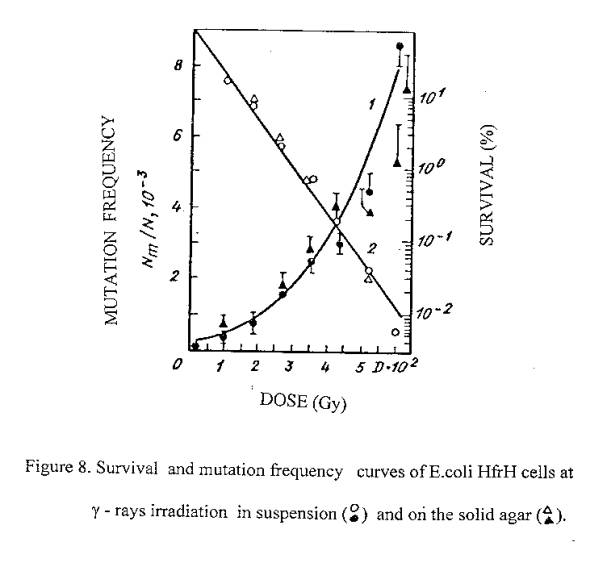
633 nm laser radiation induced mutations curve is of interesting form: in the low irradiation region the maximum of mutation frequency is observed, further the curve is reducing and at higher irradiation doses it increases again.
It is well known, that non-irradiated E.coli bacteria cells are capable of filament formation. The frequency of filament formation is depended on cells cultivation conditions. In the conditions of our experiments (grown on solid complete nutrient medium at 370C) the frequency of filament formation can be up to 30%. It was considered, that fil- gene in E.coli cells do not affect the DNA reparation processes, but increases its sensitivity to radiation (9,10).
Taking all this into account, we can suppose that the mutation frequency curve consist of two components: component of more sensitive filaments (low dose region) and component of normal cells. For the filaments, the mutation frequency curve has a shape with maximum, but for normal cells we observe linear – quadratic dependence on dose exposure (as in the case of ionizing radiation). In the favor of such suppositions the form of the cells surviving curve is also testified, which indicate heterogeneity of the irradiated culture.
These results demonstrated, that there are some general regularities of ionizing and laser radiations action on bacteria.
REFERENCES
1. Gamaleya N.F. (1977) Laser biomedical research in USSR, in Laser Application in Medicine and Biology, v.3, Wolbersht M. L., Ed. Plenum New-York.
2. Oshiro T. and Calderhead R.G. (1988) Low level laser therapy. A practical introduction. Edited by John Wily & Sons, Chichester.
3. Tokarova B., Amirtaev K.G., Krasavin E.A., Kozybek S. (1987) Express of Lac- mutations in E.coli bacteria by "sandwich seeded" method. JINR, Russia, P-19-87-1987.
4. Peck I.G., Peack M.I. (1982) Lethality in repair - proficient Escherichia coli after 365 nm ultraviolet light irradiation is dependent on fluence rate.- Photochem. Photobiol , v. 35, N3, 359 -364.
5. Raevskiy B.I. (1959) Radioactive radiation doses and its action on organism. Medgiz. p. 6 (in Russian).
6. Petin V.G., Polit P. (1969) Influence of the dose power on yeast's cells survival and recovery. Radiobiologia, v.9, N4, 492-498 (in Russian).
7. Lee D.E. (1963) Radiation action on living cells. M: Gosatomizdat. p. 287 (in Russian).
8. Kubitschek H.E., Doyle R.I. (1981) Growth delay induced in Eschericha coli by near-ultraviolet radiation: relationship to membrane transport functions. Photochem.. Photobiol., v.33, N5, 695-702.
9. Myasnik M.N. (1974) The genetic control of bacterias sensitivity. Atomizdat, Moscow (in Russian).
10. Horward - Flanders P., Simson E., Hteriot L. (1964). The excisions of thymine dimmers from DNA filament formation and sensitivity to ultraviolet in E.coli K-12. Mutat. Res., v.1, N3, 219 225.